研究紹介
統合失調症における
認知機能障害の
病態メカニズム解明
THEME 01
統合失調症は、約100人に1人が罹患する代表的な精神疾患で、幻覚や妄想などの精神病症状が目を引きますが、認知機能の低下が罹患者に共通して認められます。精神病症状が服薬によりある程度改善するのに対し、認知機能の低下に対しては効果的な治療法が確立されていません。認知機能低下は、罹患者の社会復帰を妨げる大きな要因となっています。我々は、認知機能低下の病態メカニズムを解明し、有効な治療法の解明に役立てたいと考えています。統合失調症では、知覚処理、注意、記憶などの様々な認知機能に広汎な低下が認められますが、特に多く報告されているのが作業記憶(限られた量の情報を短期間保持し、それに基づいて行動や思考を行う機能)の低下です。作業記憶は他の認知機能の基盤となっており、その低下は統合失調症の認知機能低下の中核と考えられています。作業記憶は、大脳皮質の複数の領域を含む神経ネットワークにより担われています。我々は、大脳皮質でネットワークを形成し、認知機能のための情報処理を担っている神経細胞(ニューロン)変化を解明し、認知機能低下への関与を検証しています。そのために統合失調症患者及び精神疾患の罹患歴のない対照者から提供されたヒト死後脳の解析、そして死後脳における所見を再現した遺伝子操作マウスの解析を行っています。
死後脳研究
疾患の病態メカニズムの解明には、病態が存在する器官や組織において、分子レベルの解析を行う必要があります。我々は、ヒト死後脳バンクを有するピッツバーグ大学精神医学部門との共同で死後脳研究を行っています。
大脳皮質には、興奮性のグルタミン酸を用いる錐体ニューロンと抑制性の神経伝達物質γアミノ酪酸(GABA)を用いる抑制性のニューロン(GABAニューロン)が存在します。GABAニューロンは,その形態および機能の特徴から,パルブアルブミン(PV)陽性細胞,ソマトスタチン(SST)陽性細胞,血管作動性腸管ペプチド(VIP)陽性細胞の大きく3つのグループに分けられ、それぞれが大脳皮質における情報処理に特有の機能を担っています(図1)。我々はこれまで、統合失調症の背外側前頭前野(作業記憶ネットワークの中心領域)では、GABAニューロンの中でもPVニューロンとSSTニューロンにおいて、ニューロンの機能を担う分子の発現変化があることを示してきました(図1)(Lewis et al. Nat Review Neurosci 2005, Hashimoto et al. Molecular Psychiatry 2008, Georgiev et al. Am J Psyciatry 2014)。PVニューロンとSSTニューロンは、大脳皮質において律動的神経活動(オシレーション)を形成し情報処理の効率化を担っており、統合失調症の罹患者ではオシレーションの異常が多く報告されています。すなわち、PVニューロンとSSTニューロンの機能変化が大脳皮質の機能変化と認知機能の低下に関与していることが考えられます。
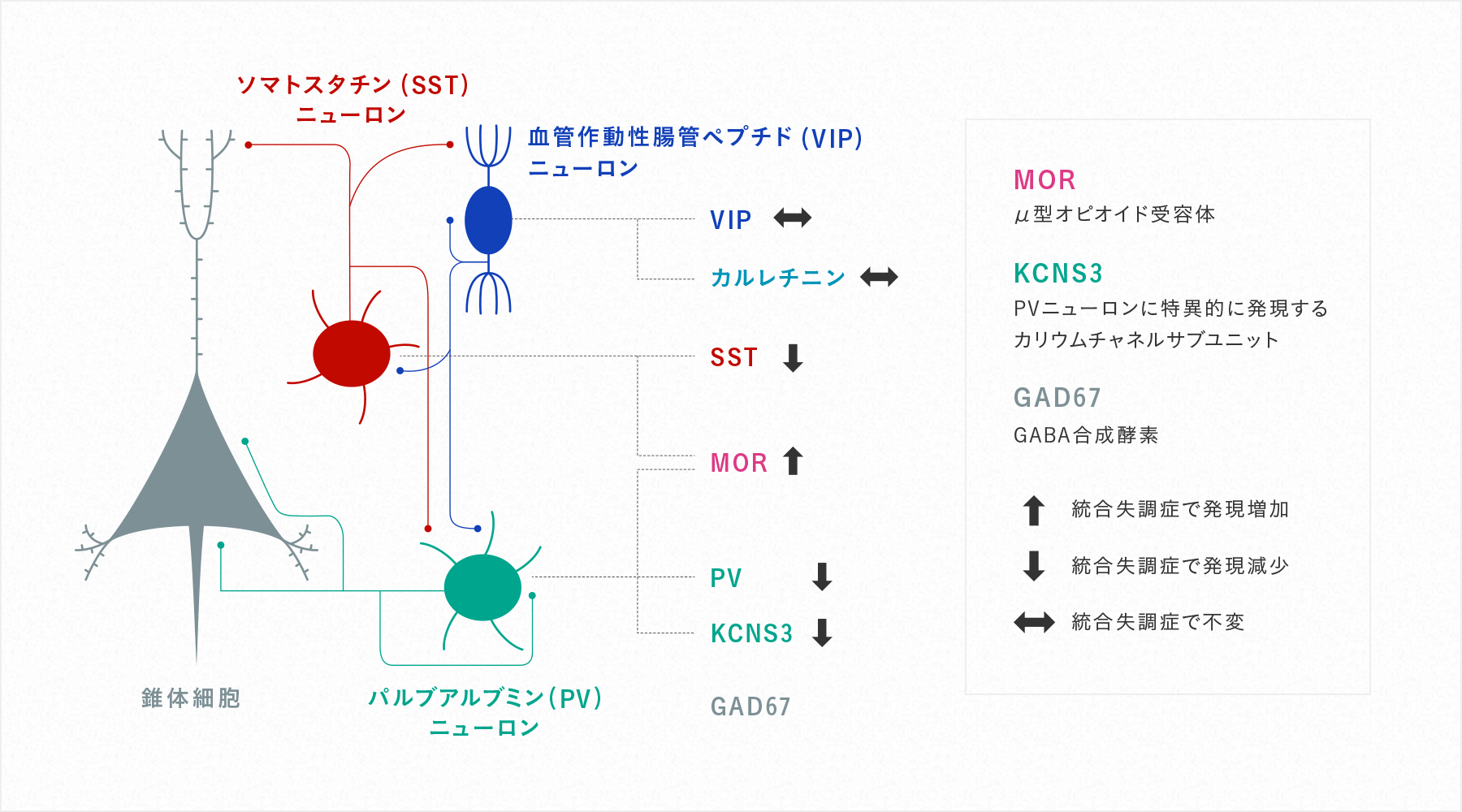
(Tsubomoto et al. Cerebral Cortex 2019より改変)統合失調症の背外側前頭前野では、PVニューロン、SSTニューロンに選択的に、その機能を担う分子の発現変化が認められる。
KCNS3遺伝子は、カリウムチャネルサブユニットをコードする遺伝子で、ヒト大脳皮質ではPVニューロンに選択的に発現し(Georgiev et al. PLosOne 2012)、このニューロンの興奮性を調節することで、オシレーションの形成に関与していると考えられます。我々は、KCNS3の遺伝子発現が、統合失調症の背外側前頭前野PVニューロンで低下していることを見出しました(Georigev et al. Am J Psychiatry 2014)。KCNS3の発現低下により、PVニューロンの興奮性が変化し、オシレーションの異常、そして作業記憶の低下につながっている可能性を想定しています(図2)。
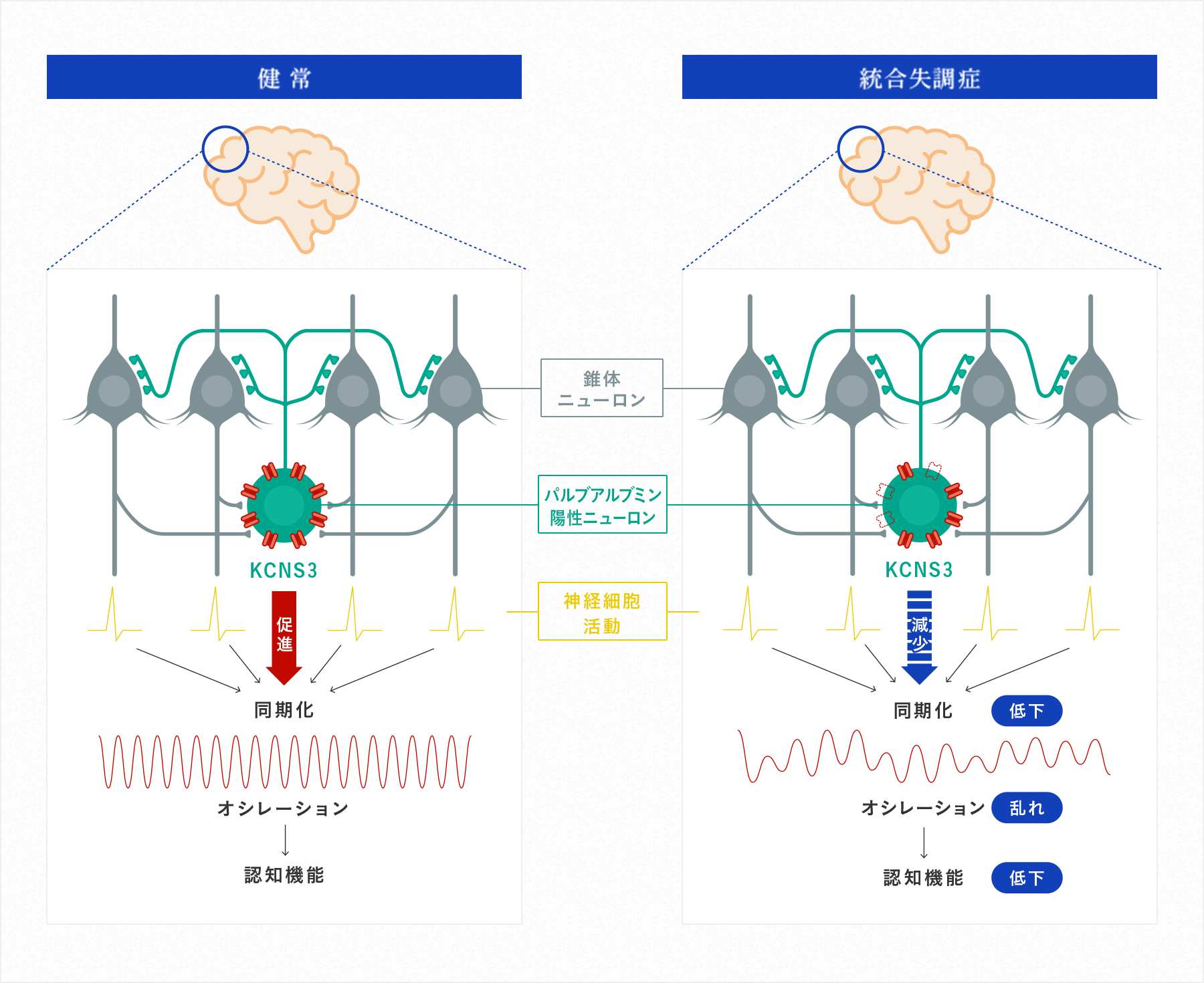
我々は、このKCNS3を含めたPVニューロンやSSTニューロンにおいてその機能を担う分子の発現変化が、背外側前頭前野だけでなく、作業記憶ネットワークを構成する後部頭頂葉や感覚野にも存在することを報告しました(Tsubomoto et al. Cerebral Cortex 2019)(図3)。また、PVニューロンとSSTニューロンの変化の上流メカニズムとして興奮性の錐体ニューロンの変化についても解析を進めています。最近、統合失調症の作業記憶ネットワーク領域において、錐体ニューロンの活動性を制御するアクチン制御遺伝子やミトコンドリア関連分子の発現変化を報告しました(Kimoto et al. Neuropsychopharmacology 2022)。
ところで統合失調症と類似の認知機能障害は、気分障害である双極性障害およびうつ病にも、統合失調症より軽度に存在し、再発性や社会適応に影響を及ぼしていることが知られています。そこで現在、図1のような介在ニューロンの変化について、統合失調症、双極性障害、うつ病の間で比較するため、対照者と各疾患の罹患者の背外側前頭前野において、PV, KCNS3, SST, VIPの発現変化を調べており、統合失調症ではPV, KCNS3, SSTに加えVIPまで発現低下が認められるのに対して、双極性障害ではPVの発現低下が、うつ病ではSSTに発現低下が認められました(Okuda et al. in preparation)。これらの結果は、統合失調症ではGABAニューロンの変化が、3つのサブタイプに及んでおり、その中でもPVニューロンの変化が著しいこと、双極性障害とうつ病ではそれぞれPVニューロンとSSTニューロンに変化が生じていることを示唆していると考えられました。
今後は、双極性障害およびうつ病の作業記憶ネットワークにおけるGABAニューロンの変化について、背外側前頭前野に加え後部頭頂野や視覚野でもこれらの特定的マーカー分子の発現を調ゆく予定です。

遺伝子改変マウス研究
ヒト死後脳で認められた所見とニューロン機能や認知機能の低下との因果関係の検証は、遺伝子操作を用いて統合失調症で認められた変化を再現したマウスを作成・解析することで行っています。例えば、統合失調症の大脳皮質PVニューロンでは、伝達物質GABAの合成酵素であるGAD67の発現が低下しており(Hashimoto et al. J Neuroscience 2003)、PVニューロンによる抑制性神経伝達の低下を示す所見として広く受け入れられています。我々は、PVニューロン特異的にGAD67遺伝子を不活化したマウスを作成し、ニューロンの興奮性の変化や行動の変化を報告しました(Georgiev et al. Schizophrenia Bulletin 2016)。現在は、KCNS3遺伝子の発現を低下させた遺伝子変異マウスを作成し、神経回路におけるPVニューロンの興奮性の解析や作業記憶の評価を行っています(図3)。最近、KCNS3の発現を正常の50%程度に減少させたPVニューロンでは、このニューロンに特徴的は高周波数(γ帯域)での規則正しい発火パターンに乱れが生じていることを報告しました(Miyamae et al. Neurobiology of Disease, 2021)。
